Chapter 25 – How Insects Shaped Biology: Darwin, Mendel, Morgan and the Modern Synthesis

We tend to think of insects as unimportant nuisances. But history reveals a much richer and sometimes pivotal role for our friends with six legs. In fact, some of the most important cornerstones of modern biology were set in place with the help of the phylum Arthropoda. In this chapter, we will look at how research on insects filled a critical hole in our knowledge base, the resolution of which led to the so-called Modern Synthesis. The latter was a massive coagulation of data that supported and reconciled Darwinian evolution with Mendelian genetics by discovering the source of variability upon which Natural Selection acted. It is, perhaps, the most important event that has taken place in biology in the last 100 years. To fully understand the magnitude of the Modern Synthesis, we need to knit together the contributions made by a number of important historical figures in biology including Charles Darwin, Gregor Mendel, and Thomas Hunt Morgan (Figure 25.1a-c).



Charles Darwin
To fully explicate the magnitude of the modern synthesis and explain the role of insects in it, we need to return to the year 1859. This was the seminal year in which a book entitled On the Origin of Species by Means of Natural Selection or the Preservation of Favored Races was published. The author, of course, was Charles Darwin (Figure 25.1a). He had been working on the book for 20 years and still might not have published it had it not been for the fact that he was about to get scooped. A much younger man, Alfred Russell Wallace, had happened upon much the same theory while recovering from the ravages of malaria in Indonesia. Wallace sent a letter to Darwin with a short manuscript entitled On the Tendency of Varieties to Depart Indefinitely From the Original Type. Darwin, himself, realized that Wallace had restated Darwin’s theory of natural selection without using the term and in much shorter form. A bit panicked, Darwin consulted his good friends Charles Lyell and Joseph Hooker who convinced Darwin to jointly publish his 800-page book alongside Wallace’s short essay. The idea that species could change through the agency of Natural Selection was, thus, published.
Darwin’s basic idea about how species change was fairly simple. His first premise is something that we would take as given today, namely, that there exists variation in nature. You have only to look at the members of your own family to verify this. If you have siblings with the same biological parents, it will be readily apparent that overall look different from your siblings, unless you are an identical twin of course. The second part of Darwin’s argument was that the variation observed was heritable. Today, we would say that it has a genetic basis, and again, this is obvious: you share more physical features with your parents and siblings than you do with people who are not closely-related to you. Third, Darwin believed that there are more offspring born than can be supported by the environment. In other words, there is a chronic mismatch between the number of mouths to feed and the available resources. This led to a “struggle for existence”—not a phrase coined by Darwin, but one that is often used to describe this situation. The fourth element of Darwin’s thesis was that organisms possessing favorable variation (those that were best-suited for the present environment) would produce more offspring because they were better equipped to acquire limited resources. Finally, over long periods of geological time, organisms would accumulate several small changes that could lead to major differences in the appearance and traits of organisms, creating new species.
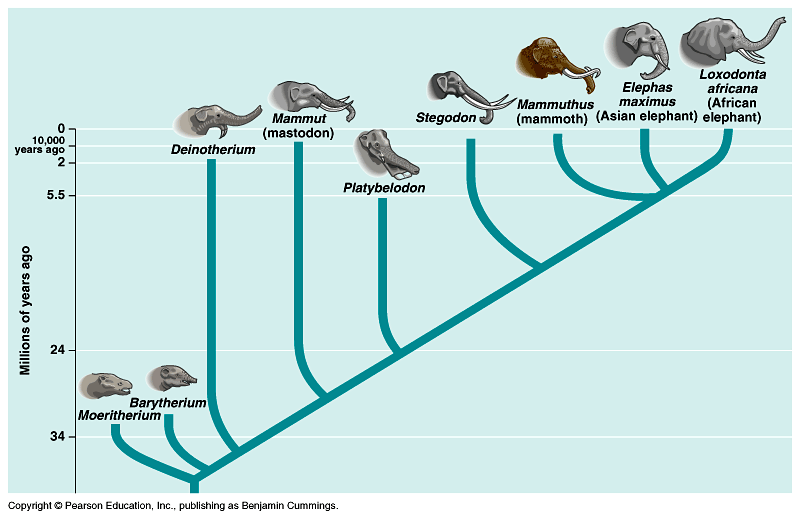
Ever the reductionists, we sometimes contract Darwin’s lengthy argument and supporting evidence into the idea that the variation of life on earth comes from Descent with Modification and that the mechanism for accomplishing the modification is Natural Selection. Descent with Modification results in the kind of family tree that you see presented in Figure 25.2 (click on link) where every node (intersection of two lines) represents the diversification of new types of organisms from ancestral types so that there is enough of a difference to prevent breeding between the ancestral and new types. When this happens, speciation has occurred. As an aside, the biological definition of species is that species are members of a population that are actually or potentially capable of interbreeding and producing fertile offspring.
{kind=link}
Gregor Mendel
We now move to the second protagonist in our tale. Genetics and the mechanism of heredity were actually worked out by a contemporary of Darwin’s, Gregor Mendel (Figure 25.1b). Mendel was an Austrian monk who floundered as a farmer and eventually entered a monastery. He experienced several more failures, such as twice failing the exam to become a teacher. Even when he settled on performing biological experiments in the garden of the monastery where he lived, his path was not simple. Mendel’s initial choice of test subjects, mice and bees, proved disastrous. But, eventually, he turned his attention to pea plants and that turned out to be a fateful decision. Indeed, he deduced the fundamental laws of genetics and became the Father of Genetics as a result. Simply stated, these laws are as follows:
1) The Law of Segregation
2) The Law of Independent Assortment.
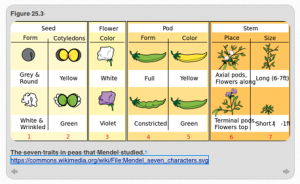
Let’s take a look at how these work. First, we need to appreciate the genius of Mendel’s choice of pea plants. He focused on seven traits that involved flower color, seed shape, and plant size (Figure 25.3). We cannot tell if these choices were fortuitous or whether he could tell that these traits behaved in a “reliable” way that made them easier to study. Either way, this choice of both pea plants and the seven particular traits he decided to follow in his breeding studies made all the difference. The traits Mendel selected positioned him for success because each trait had two forms (alleles), which had a simple relationship of dominance to recessiveness. If this had not been the case, Mendel might never have deduced the laws of genetics.
Mendel began doing crosses among pea plants having different characteristics. One of the things that he learned very quickly is that when parents differed in a trait, the offspring show one trait or the other, not a mixture of both. This meant that the reigning theory, the so-called blending theory of inheritance was wrong. The idea here was that when two parents differ in a trait, such as when you breed a black cat to a white cat, the outcome is sort of halfway between the two parents or gray. Mendel showed that that was wrong.
Mendel also provided some insight into other mechanics of heredity. His law of segregation, which we will look at in a moment, helped us to understand that each parent, male and female, contributes equal amounts of genetic material to the offspring.
Finally, Mendel demonstrated that the genes for different traits behave independently from one another across multiple generations. This became known as the Law of Independent Assortment. How did Mendel do all of this? First, he observed simple patterns that occurred in crosses—such as dominance and recessiveness. Mendel learned that dominant genes are so called because when a dominant gene is present, it will mask the appearance of the recessive gene. You know this from your own life. Assume that only one gene is responsible for hair color in humans (actually, it is 4 genes, each with 2 different variants or alleles) and that the gene for brown hair in humans is dominant to the gene for blond hair. If we represent allele brown hair is represented with a capital “B” and the allele for blond with a small “b”, we can easily demonstrate how Mendelian inheritance works.
Remember that both parents contribute equally to the offspring. Therefore, when both parents contribute an allele for brown hair (“B”) then the baby will be “BB” and will have brown hair However, if one parent gives baby a “B” allele and the other parent gives the baby an allele for blonde hair (“b”), then baby will be “Bb,” and will have brown hair. This is because the “B” allele is dominant to the “b” allele and masks the expression of “b.” As a result, the only way you can get a blond-haired baby is if both parents contribute a “b” allele to their offspring. If both parents contribute a “b” allele to the baby, then the baby will be “bb” and will have blond hair. In other words, two copies of the “b” allele must be present in order to have blond hair and this is one reason why blond hair is less common than brown hair.
Before we go on, let’s take a moment to review a few terms (refer back to Sidebar 14.1 for more detailed information). When we refer to the actual genes an organism has, we are talking about its genotype. There were three genotypes in the human hair example which were: “BB,” “Bb” and “bb.” However, when we are talking about what an organism looks like, we are referring to its phenotype. For the example we used, there are only two phenotypes. This is because both “BB” and “Bb” will result in brown hair while only “bb” will produce blond hair.
Another set of terms we need to introduce are homozygous and heterozygous. Homozygous means that an organism has to copies of the same gene, i.e., “BB” or “bb.” In contrast, a heterozygous individual has mixed genes or one copy of both genes or is “Bb” in this example. We can also mix terms like homozygous dominant (“BB”) or homozygous recessive (“bb”).
Mendel’s test subject, the pea plant was operating on the same system. Mendel was able to deduce what was going on by noticing the resurgence of particular patterns in the offspring from his crosses. He noticed two general types of patterns: sometimes in his crosses he had true-breeding plants that always produced offspring that looked just like their parents. Other times, he would cross two plants that had a dominant phenotype, and approximately 75% of their offspring would have the dominant phenotype and 25% would have the recessive phenotype.
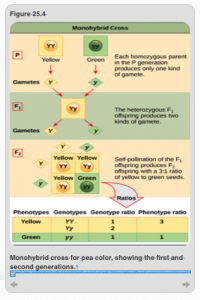
Mendel figured it out using a monohybrid cross (Figure 25.4). To demonstrate this concept, we could take any one of the seven traits he studied. This example involves the seed color of the peas of which there were two states: yellow or green. These are two phenotypes. Here we will represent the yellow trait with a big “Y” a little “y” for the green trait. In this experiment, Mendel was working with what are called true-breeding types. That is, if you cross a yellow-looking parent with another yellow looking parent, you will always get 100% yellow offspring. Similarly, if you cross a green seeded-parent with another green-seeded parent, you will always offspring that produce greed seeds. Another way of expressing the idea of true-breeding is that all of the parents are homozygous (“YY” for true-breeding yellow; “yy” for true-breeding green).
With that information, you should be able to understand the cross in Figure 25.4. Mendel crossed one true breeding homozygous dominant parent, “YY,” with a true breeding homozygous recessive parent, “yy.” What would the outcome of this cross be?
Law of Segregation
The easiest way to understand this is to set up what geneticists call a Punnett Square, a simple version of which is found at the bottom of Figure 25.4. The alleles of parent 1 are labeled on the top of the table and the genes of parent 2 along the side. The answer is provide in the figure: the “YY” parent is only capable of producing gametes (sex cells) that have one copy of the “Y” allele. The second parent’s genotype is “yy” and is only capable of producing gametes that have a “y” allele. When the gametes of the two parents come together, all of the offspring will, therefore, be “Yy” heterozygotes that will have a yellow phenotype. In other words, 100% of the offspring in the first filial generation (F1) will be yellow. This is one of the two patterns that Mendel saw repeatedly.
The second consistent pattern emerged when he took two of the offspring from the F1 and bred them in a second cross, making the second filial generation (F2; bottom half of Figure 25.4). You already know the genotypes of the two parents: they are both heterozygotes having one copy of the “Y” allele and one copy of the “y” allele. Next, we need to identify the gametes that both parents produce. Since they are heterozygotes, both parents can produce gametes that have the “Y” or “y” alleles. To determine the identity of the potential offspring, all you have to do is fill in the boxes by simply doing the cross and entering the correct alleles from parents 1 and 2 for each box.
If you set this up and do the crosses correctly, there will be three genotypes: “YY,” “Yy,” and “yy.” Just as before, there will be two phenotypes: yellow and green. If you look at the Punnett square in Figure 25.4, you will see that there is one individual with the homozygous dominant “YY” genotype, 2 individuals with the heterozygous “Yy” genotype, and one individual with the homozygous recessive genotype “yy.” This distribution of genotypes explains why Mendel observed a 3:1 ratio of dominant vs. recessive phenotypes in the offspring when he did this type of cross. This observation allowed Mendel to uncover the first of his two laws of hereditary: the Law of Segregation. That is, alleles segregate onto gametes (sperm or eggs). This allows recessive alleles to apparently disappear as they did in the first cross, but to segregate from the dominant allele when the organism was producing gametes so that the recessive trait could reappear in subsequent generations as they did in the second cross.
Law of Independent Assortment
Mendel’s second law, the Law of Independent Assortment, can only be understood in the when examining the inheritance of two or more genes such as seed color and seed texture. This law states that during the sorting of alleles that occurs at reproduction, or the parceling of alleles into gametes, occurs randomly and all possible genotypes will be equally represented among the gametes. If we are working with two genes (a dihybrid cross), both of which exist in the heterozygous condition, then each parent could produce gametes with one of 4 possibilities. Let’s say that first trait we are now looking at are the color of the pod. In this case, the pod is either green, represented with a “R” allele because it is dominant, or yellow and represented with a little “r” allele because it is recessive. Then the second trait we are considering is whether the pods are constricted, represented with an “Y” because this is the dominant trait, or full, represented with an “y” because it is recessive. Thanks to the Law of Independent Assortment, the two genes and the alleles therefrom assort independently of one another and can therefore produce all possible phenotypes in the potential offspring. Therefore, if a parent is heterozygous for both traits (“RrYy”), that parent can produce gametes with one of 4 different combinations of these alleles: “ry,” “RY,” rY,” or “RY.”
As you will see in Figure 25.5, if both parents are heterozygous for both traits, there are eight possible genotypes (and multiple ways to get some genotypes) and four possible phenotypes that exist in a 9:3:3:1 ratio (9 will be green and have constricted pods, 3 will be green with full pods; 3 will be yellow with constricted pods, and 1 will be yellow with full pods). Again, this shows that heterozygous parents can produce offspring with all possible combinations of phenotypes for these two traits. This example uses only two genes. The complexity grows exponentially as additional genes are added to the mixture but as this is not a genetics text, we will spare you the details.

Figure 25.5: Results of a dihybrid cross showing Mendelian’s Law of Segregation and Law of Independent Assortment. Here the traits are whether the pea pods are green (R) or yellow (r) and whether the pod shape is constricted (Y) or full (y). Parents that are heterozygous for both traits produce gametes with one of 4 combinations of the traits in equal numbers. The genotypes and phenotypes of their offspring are shown in the resulting Punnett square. This image is has been released into the Public Domain by Lady of Hats.
{kind=link}
This is how Mendel deduced the laws of inheritance via a highly experimental and mathematical demonstration of how nature worked. You, too, should be able to explain how Mendel arrived at his conclusions with crosses involving single traits. Maybe it will help you can appreciate the difficulty of operating in the dark without key pieces of information such as how reproduction works and how gametes are formed biologically.
Seeming inconsistencies between Darwin and Mendel
Unfortunately, Mendel’s important and amazing discoveries were largely ignored during his lifetime. He, at one point, sent a copy of his work to Darwin, who apparently never bothered to read it, in large part because Darwin could not read German. Which was really too bad because other scientists critiqued Darwin’s work for lacking a mechanism explaining how traits could be inherited. It wouldn’t be until 1900 when three scientists (de Vries, Correns, and Tschermak) simultaneously re-discovered Mendel’s work and understood its importance and relevance to the fledgling field of genetics. Mendel solved the mechanism of inheritance but neither Mendel’s work, nor anyone else at the time could explain where the variation came from; how is it that peas could be yellow or green, pods could be constricted or full? Without that information, it was still not possible to explain speciation.
Therefore, in the early 1900s, scientists were divided into two camps due to apparent inconsistencies between Mendelian genetics and Darwin’s Theory of Natural Selection to explain how new species could arise. Darwin’s Theory stated that new species arose by accumulating several small changes over large periods of time. Yet, Mendel explained how discrete differences in traits could be inherited and re-appear, and among his pea plants there weren’t any intermediate forms (a plant that produced green seeds when crossed with another that produced yellow seeds produced offspring with either green or yellow seeds; not greenish yellow). How could these be reconciled?
Biologists belonging to he biometric school, were proponents of natural selection and rejected Mendelian genetics as they claimed that the simple mechanism proposed could not explain the continuous variation that we observe in real populations today (e.g. people show all ranges of skin color and height; we are not either black or white, short or tall).
In contrast, supporters of Mendel’s work fell into the saltationist school which proposed that new species proceeded by big jumps rather than undergoing lots of intermediate forms. One prominant researcher in this camp, Hugo deVries, proposed the theory of mutation, as a mechanism by which a new species could be generated rapidly.
As it turns out, the theory of mutation was wrong, but was a significant step forward at the time. The idea here was that the genetic material, referred to as pangenes, underwent alterations which were called mutations. This is actually true. The part that they got wrong was that they believed that pangenes were genetic material produced by all parts of the body. The pangenes then traveled to the gonads where they were taken up by sex cells and that’s how they could be passed on to the next generation. The other part that they got wrong is that a new species cannot be created by acquiring one or even several mutations all at once.
Somehow, there needed to be an experimental resolution of which idea was correct. And here, as was the case with Mendel, the critical element of the solution was finding the right test organism. As we have already learned, Mendel was either extremely lucky or incredibly astute in choosing pea plants because he was able to work with an organism that had easy-to-follow traits that showed simple dominance and recessiveness. In truth, genetics is often much more complicated than that and, when you are trying to work out the fundamentals (as opposed to elaborating on established principals), it is absolutely critical to have a cooperative (read: simple) test organism to help you out.
Thomas Hunt Morgan
And help was on the way. Here, at last, we get to why insects are relevant to this quest. In the laboratory of Thomas Hunt Morgan (Figure 25.1c), a scientist who had done considerable work with de Vries, the answer was lurking. Like de Vries, Morgan had started out as a mutationist but thought de Vries had gotten key parts of the theory wrong. Morgan benefitted from his insistence that we needed to learn about mutation and its role in heredity through experimental studies. And Morgan wanted to show that the production of mutations could, in fact, produce new species. In order to do this, he was making huge strides using Drosophila melanogaster, otherwise known as the fruit fly, as a test organism.
Morgan began working with fruit flies and subjecting them to radiation and toxic chemicals in order to induce mutation—a theme rapidly adopted and expanded upon by so many science fiction movies to come. It is said that part of Morgan’s success was due to his uncanny ability to pick out fruit flies form a swarm of hundreds that sort of looked different. With his super eyesight, he found the “white-eyed male” in 1910. He then, like Mendel, began doing breeding experiments. When he bred the white-eyed male back to his red-eyed (normal) sisters, he found that all the resulting offspring had red eyes. But, if he selected from the offspring a red-eyed female and a red-eyed male and crossed them, he got a 3:1 ratio of red to white eyes in the second generation.
So far, it sounds like a cross between two heterozygotes, right? Well, it’s actually a little more complicated than that. You see, it was only the male offspring that ever had white eyes, never a female offspring. And recall that the cross between the white-eyed male and red-eyed female all of the offspring, male and female, had red eyes.
Figure 25.6 gives a graphic representation of Morgan’s experimental results. And now we need to do a little back-peddling to explain a weird thing that is going on. It’s called sex-linked inheritance (see Sidebar 25.1 for more information on module page). Like humans, female fruit flies have two X chromosomes. And males have an X and a Y chromosome. So, if a particular gene is found only on the X chromosome, it means that female fruit flies will have two copies of that gene while male fruit flies will have only one copy of the gene, found on their single X chromosome. And, if the gene has two forms (alleles) of the gene (“w+” = red eyes; “w” = white eyes) which, like Mendel’s pea plants, show simple dominance and recessiveness, then a female fruit fly with two copies of the gene, could have three potential genotypes (“w+ w+”, “w+ w”, “ww”) and two phenotypes (red eye, white eye). A male fruit fly with only one copy of the gene can have two genotypes (“w+ Y” or “wY”) and two phenotypes (red, white). Importantly, the male with one copy of the gene will show the recessive phenotype with one copy of the gene. Remember, in Mendel’s experiments, the peas had to be homozygous recessive to show the recessive trait. For sex-linked traits, females do need to have two copies of recessive traits for them to be observed. However, males do not and will show the recessive trait with only a single copy.

Figure 25.6: Experimental cross performed by Thomas Hunt Morgan, illustrating the X-lined inheritance of white-eyed mutation in fruit flies. Results from reciprocal crosses between red-eyed (Red) and white-eyed (White) Drosophila in Morgan’s experiments. w + = red allele (wild-type allele) and w = white (mutant allele). In sex-linked inheritance, alleles on sex chromosomes (XY) are inherited in predictable patterns on Drosophila sex-chromosomes. Image by Yasmine Mrabet. GFDL, CC-BY-SA
{kind=link}
At the top of Figure 25.6, we see the normal red eyed female has two X chromosomes and two copies of the normal gene (“w+ w+”). She is mated to a mutant, white-eyed male that has one copy of the recessive allele (w) which is found on his X chromosome. He also has Y chromosome with no gene for eye color. Since the female can only produce the gene for eye color, all of her offspring, both male and female, will receive one copy of the “w+” gene from her. The male has the white eye allele or “w” on this X chromosome which he will contribute to his daughters who are XX and will, therefore, receive an X chromosome from each parent. Since all of the daughters from this pairing will be “w+ w”, all of the daughters will have red eyes but will be carriers of the white-eye trait. Male offspring from this cross will receive an X chromosome from their mother with the red-eyed allele (“w+”) and, thus, will be red-eyed. Males will receive the Y chromosome from their father, but since the Y chromosome lacks the gene for eye color, there will be no modification of eye color from the male parent. The outcome of this cross is detailed in a Punnett Square in Figure 25.6 where the genotypes of the parents are explicitly connected to the sex chromosomes of each parent.
But in a second cross (F2 on the right hand side of Figure 25.6) with a red eyed female carrier from cross (“w+ w”) with a normal red eyed male (“w+ Y”), things start to get interesting. The female parent is a heterozygote who can contribute either an “w+” or “w” allele to her progeny. In contrast, the male has only the “w+” allele, so that is all he can give to his progeny. We will track the X chromosome so that we can determine sex and, thus, the phenotype of the progeny. Here, half of the female progeny will have two copies of “w+” and will be normal, having red eyes. The other half of the females will be carriers for “w” and will be heterozygotes (“w+ w”) and will have red eyes just like their “w+ w+” sisters. But, among the male progeny, half will have gotten a normal “w+” allele from mom and will have red eyes and the other half will have gotten the “w” allele from mom and will have be white eyes. This results in a 3:1 ratio of red to white eyes in the second generation (F2). This arises because all of the daughters have at least one copy of the “w+” allele that causes red eyes. In contrast, 50% of the male offspring receive an “w+” gene from their mother with the other 50% receive the recessive “w” allele from the mother. Thus, half of the male progeny will have red eyes and the other half will have white eyes. This would not happen if the relevant gene were not on the X chromosome and inheritance of the eye color gene were not sex-linked.
Morgan’s discovery of sex-linked inheritance opened up other avenues of investigation. Another trait which behaves the same way in Drosophila is body color, where the options here were grey (normal) and yellow (mutant). Unlike traits that Mendel had examined in pea plants like pod color and shape, the alleles for eye color gene and body color were almost always inherited together. This means that these two traits did NOT segregate independently onto separate gametes as Mendel’s second law stated. It turns out that this is because the two genes are found on the same chromosome (X), and these genes are located very close together. Because of their close physical proximity, the two traits tend to be stuck together and move onto the same sperm or egg cell. This finding was instrumental in showing that genes are located on chromosomes, and not some other component of sperm or eggs. But, most importantly, thanks to Morgan’s work, mutation was now understood to be the source of genetic variation in nature.
Chapter Summary
And with the work of these three scientists, Darwin, Mendel, and Morgan, the revolution in biology called the Modern Synthesis became possible. Morgan induced genetic changes (changes in the structure of DNA) using X-rays and toxic chemicals to disrupt DNA structure. Thanks to this work, he was able to generate several new variants in fruit flies, and this revealed that changes in DNA structure are how differences in genetic diversity arise. But contrary to what Morgan initially thought, he never created a new species of fruit fly by a single or even mutations.
Morgan’s work therefore brought together two schools of thought that were diverging from one another. His studies showed how Darwin’s ideas about genetic variation were related to Mendel’s laws of inheritance and that, contrary to what scientists thought in the early 1900s, both Darwinism and Mendel were compatible. Mutations were the source of variation, that could be inherited and acted upon by natural selection. Over time, accumulating multiple changes, new species could be generated. Further, Morgan showed that mutation did not result in speciation in a single step suggesting that Darwin was correct; instead new species could only be generated from the cumulative effect of extensive adaptive radiation driven by natural selection.
One more monumental discovery was needed to finish the script for the Modern Synthesis and that came from Watson and Crick who discovered the three-dimensional structure of DNA (Deoxyribose Nucleic Acid). The latter served the important function of tying all of the loose strands together into a coherent model. The Model of Watson and Crick set the platform for understanding how DNA, and consequently heredity, functioned: how DNA stored genetic information; how that information was precisely copied and packed into cells; how disruptions in the sequence of DNA could result in mutation; how mutation could either result in a functional alternative gene or a complete disaster. With Darwin’s focus on heritable variation, Mendels’ work on how traits were determined from genes, Morgan’s work on mutation as the source of variation and Watson and Crick’s work on DNA structure, the picture was complete. As is true of all good theories, it provided great explanatory and predictive power which has driven the course of biological research ever since (Figure 25.7).

Figure 25.7: Several major ideas about evolution came together in the population genetics of the early 20th century to form the modern synthesis, including genetic varitaion, natural selection, and Mendelian inheritance. This ended the eclipse of Darwinism and supplanted a variety of non-Darwinian theories of evolution that were present in the early 1900s. Image by Ian Alexander CC BY-SA 4.0
Among the big ideas explained or correctly predicted, we have the following:
• We now have a mechanisms for understanding why there is so much biodiversity on our planet.
• We now understand both how it is possible for you look like your parents or not look like your parents.
• We understand that natural selection works at the level of genes but within populations, not upon individual organisms as Darwin originally supposed.
• We have a better understanding of how new species can be formed from accumulated change AND by other mechanisms as well.
Among those other mechanisms, we have genetic drift, a variety of reproductive isolating mechanisms, as well as interaction of the genome with viruses and other contributors of DNA to cells.
Is it right to attribute all of this to the fruit fly? Well, maybe not explicitly. But even Darwin’s work depended heavily on the examples that insects provided to biodiversity. The information provided by Morgan was critical to securing the Modern Synthesis and it, clearly, benefited from the insect model that he developed. Could it have arisen with some other test organism? Yes, probably. But insects offered some huge advantages like their short generation time, high fecundity, ease of handling and cultivation, and their susceptibility to mutation. The ease of finding mutants that could be distinguished by easily discernable traits like eye and body color was also a critical element. In genetic research, the challenge has always been to find the right test organism. There is no doubt that the use of the fruit fly made it easier for Morgan to figure out how mutations worked. It could have happened with another organism but it almost certainly would have taken longer. We are very lucky that the fruit fly was there to speed our understanding.
References
Chapter 25 Cover Photo: Drosophila melanogaster. Public Domain: Ktbn. Accessed via commons.wikimedia.org
Figure 25.1a: Charles Darwin. CC0 Public Domain: Corbis Images. Accessed via https://commons.wikimedia.org/wiki/File:Charles_Darwin_1880.jpg
{kind=link}
Figure 25.1b: Gregor Mendel. CC-BY 4.0: Hugo Iltis. Accessed via https://commons.wikimedia.org/wiki/File:Gregor_Mendel_oval.jpg
{kind=link}
Figure 25.1c: Thomas Hunt Morgan. CC0 Public Domain: Johns Hopkins yearbook, 1891. Accessed via https://upload.wikimedia.org/wikipedia/commons/8/8f/Thomas_Hunt_Morgan.jpg
{kind=link}
Figure 25.2: Descent with modification. Copyright: Pearson Education, Inc. publishing as Benjamin Cummings. Accessed via http://www.bio.utexas.edu/faculty/sjasper/bio212/histevolth.html
Figure 25.3: Mendel’s pea traits. CC0 Public Domain: Mariana Ruiz (LadyofHats). Accessed via https://commons.wikimedia.org/wiki/File:Mendel_seven_characters.svg
{kind=link}
Figure 25.4: Monohybrid cross. CC-BY-SA 4.0: Boundless. “The Punnett Square Approach for a Monohybrid Cross.” Boundless Biology Boundless, 08 Aug. 2016. Retrieved 10 Mar. 2017 from https://www.boundless.com/biology/textbooks/boundless-biology-textbook/mendel-s-experiments-and-heredity-12/patterns-of-inheritance-95/the-punnett-square-approach-for-a-monohybrid-cross-415-11642/
Figure 25.5: Dihybrid cross. CC-BY-SA 3.0: Yahadziia. Accessed via https://en.wikipedia.org/wiki/File:Grasak.JPG
{kind=link}
Figure 25.6: Fruit fly eye color inheritance. Copyright: The McGraw-Hill Companies, Inc. Available at https://online.science.psu.edu/biol011_sandbox_7239/node/7394
Figure 25.7: Punnett square for fruit fly eye color. By Chrissy Spencer, adapted from OpenStax Biology. Available at http://bio1510.biology.gatech.edu/module-4-genes-and-genomes/4-4-linkage-sex-linkage-and-pedigree-analysis/
Additional Readings
Allchin, D. (1997). Thomas Hunt Morgan and the White-Eyed Mutant. In Doing Biology. Pearson Education Limited.
Kohler, R. (1994). Lords of the Fly. University of Chicago Press.
Morgan, T.H. (1910). Sex-linked Inheritance in Drosophila. Science: 32: 120-122.