Chapter 2 – The Breath of Life in Insects and Humans

As you are reading this chapter, you will inhale several times per minute and, in so doing, draw air into your lungs. Fortunately, you don’t have to remember to do this. If you did, at some point in your life, you might have forgotten to inhale and you would no longer be with us. The task of breathing (thankfully) has been turned over to your autonomic nervous system which takes matters out of your conscious control and makes sure that you breathe—even when you’re sleeping and can’t think about it.
However, have you ever thought about why insects and humans have to breathe? The simple answer is to get oxygen. But why do we need oxygen? That is a complicated physiological issue that we won’t pursue in great detail. Suffice it to say, you take atmospheric oxygen (O2) into your lungs so that your circulatory system can extract oxygen from the air, bind it to hemoglobin in your blood, and distribute the oxygen to every cell in your body. In your cells, oxygen serves as a terminal electron acceptor at the very end of the electron transport chain in your mitochondria. Yes, we know you don’t really want the details, but without oxygen in your cells doing its electron-accepting function, your cells wouldn’t be able to produce enough energy to run a big organism like you. Your cells can produce small amounts of energy without oxygen, but the byproduct is lactic acid, which is why you can literally “feel the burn” during intensive periods of exercise. To avoid this unpleasant sensation, we use oxygen to ramp up our ability to produce massive amounts of usable biochemical energy from the food we eat.
Insects need oxygen in their cells for the same reason and, therefore, they too take in atmospheric oxygen to distribute to all of their cells. But wait, I hear you cry: “Insects don’t have lungs!” That is certainly true. But insects and humans have roughly the same problem: we both need to get oxygen out of the air and into our cells, and also distribute nutrients, hormones, and immune cells throughout our bodies. However, the structures and processes we use to do this are quite different. In this chapter, we will compare and contrast the respiratory and circulatory systems that allow humans and insects to complete these functions.
Eukaryotic and Multi-Cellular Origins
For three billion years of life’s history on planet earth, the only extant (surviving) life forms were individual cells. But they were nothing to sneeze at. After all, they were the product of another couple of billion years of endless testing and experimentation until, at last, a single cellular organism with a template for its own replication evolved. For three billion years, endless variants of the single cell were produced, which successfully invaded nearly every environment on the planet, from unbelievably hot sulfur vents at the bottoms of our oceans to the frigid depths of Antarctic lakes. And as we mentioned in the previous chapter, two cells from separate domains of life, a large archaeal cell and a small bacterial cell, combined to form an entirely new single-cellular organism: a eukaryotic cell that had a true nucleus enclosing its DNA. These new eukaryotic cells were larger and more energy efficient than their prokaryotic ancestors, but were less metabolically flexible in that all of them required oxygen. However, as long as they had access to oxygen, these eukaryotic cells thrived and eventually were able to form larger and more complex multicellular organisms.
As with everything in biology, there are both benefits and challenges to being multicellular. On the one hand, multicellularity allowed different cell-types to specialize into specific tasks, such as the epidermal/skin cells that protect the organism or neural cells that convert and transmit chemical and electrical impulses to one another. On the other hand, multicellularity imposed new challenges, perhaps the greatest being how to deliver oxygen to each cell. Eukaryotic cells can survive for limited periods of time without oxygen via processes such as lactic acid fermentation, mentioned earlier. And some single-celled eukaryotic organisms can go their entire lives without oxygen like Brewer’s yeast, which uses alcoholic fermentation to produce energy in the absence of oxygen (and make delicious beer, wine, and other spirits in the process). However, if any cell in a multicellular eukaryotic organism is deprived of oxygen for an extended period of time, it will die.
Why is this? As you will remember from Chapter 1, the smaller, invading bacterial cell component of eukaryotic cells were specialized in aerobic metabolism that required oxygen. These small bacterial cells provisioned their archaeal host cells with large amounts of ATP (adenosine triphosphate), the energy currency that all cells use for every task. This increase in ATP was fueled by the addition of a new metabolic scheme called the Kreb’s Cycle, as well as the electron transport chain. Both the Kreb’s cycle and the electron transport chain required oxygen to work, but this system was able to crank out many times the ATP that was possible when only anaerobic (i.e. no oxygen) metabolism was present. Both of these processes were provided by the internalized bacterial cell. Over time, the association between the bacterial cell and its host cell became closer and closer until neither could live without the other; the eukaryotic host became completely dependent on its bacterial partner to meet all of its energy needs, and in turn the bacterial cell lost more and more of its DNA, and with it, its ability to make proteins or support itself independently. Eventually the internal bacterial cell became the mitochondria, or cellular power factories, that are present in nearly every cell of every eukaryotic organism. Thus, thanks to highly efficient and productive mitochondria, clusters of eukaryotic cells were able to form and generate enough energy to support each cell, but only if each cell had access to oxygen.
Although larger groups of cells could share more energy and resources among themselves and specialize to complete diverse tasks, size was still a limiting factor. This is because such organisms lacked a mechanism for getting oxygen to each cell. Consequently, the earliest multicellular eukaryotes, such as slime molds and other small organisms, were constrained by their ability to transport oxygen among the cells, and remained quite small. A massive expansion and diversification of organismal types on the planet was not possible until a variety of additional adaptations took place. Such adaptations provided living organisms with a respiratory system for drawing oxygen into very large bodies and a circulatory system for distributing the oxygen, and the resulting ATP produced, around a body comprised of millions of cells. With each new adaptation, the size of organism that could be sustained increased, resulting in the immense diversity of multicellular organisms on earth today.
Human Respiratory and Circulatory Systems
Before discussing how insects overcame the difficulty of distributing oxygen and nutrients to their cells, we will start by discussing respiratory and circulatory systems that are much more familiar to you: our own. In humans, the respiratory (i.e. oxygen delivery) and circulatory (i.e. distribution) systems are inextricably linked. We draw air in through the nasal cavity, after which, oxygen-laden air moves into the lungs as shown in Figure 2.1.
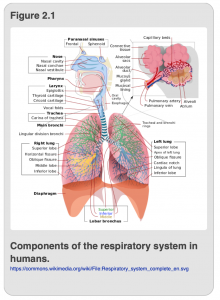
Once the air reaches the lung, oxygen is concentrated by diffusing into blood vessels and binding to a special protein called hemoglobin. The blood is now oxygenated and begins its journey through the rest of the body, courtesy of the heart, whose pumping activities direct the oxygenated blood into the aorta for distribution. Oxygenated blood moves from the aorta into smaller arteries and then into even smaller capillaries which bring the oxygenated blood into proximity with the waiting cells throughout the body.
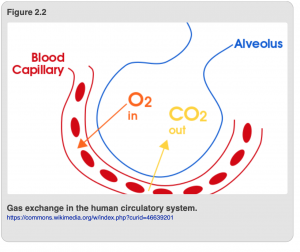
Another form of gas transfer takes place at cellular level too (Figure 2.2). Specifically, carbon dioxide (CO2), a waste product produced by cells as a result of running cellular processes, diffuses into capillaries where it also binds to hemoglobin in the blood. The deoxygenated blood then moves into veins, which convey the carbon dioxide-laden blood back to the lungs via the pulmonary artery. Within the lung, the pulmonary artery branches into smaller and smaller capillaries which run through specialized chambers called alveoli. Within a single alveolus, carbon dioxide and oxygen diffuse down their concentration gradients such that carbon dioxide moves out of the blood and into the lung, and oxygen moves out of the lung and into the blood. The carbon dioxide is removed from your lungs as you exhale each breath. In summary, it is oxygen in, carbon dioxide out as a result of the combined activities of the circulatory and respiratory systems.
Now for a few more salient details about the circulatory system in humans. First, because the blood is contained in vessels (arteries, veins, and capillaries), the circulatory system in humans considered to be a closed circulatory system. Another important point is that, in addition to providing a means to exchange oxygen and carbon dioxide, the circulatory system also has other critical functions. It circulates a variety of critical nutrients, hormones and immune cells around the body which are distributed to cells throughout the body. Therefore, the circulatory system plays a lead role in immune responses and the distribution of hormones around the body.
Insect Respiratory System
What about insects? As previously mentioned, insects have to take in oxygen to circulate to their cells and expel carbon dioxide, just as humans do. But, insects don’t have lungs, so how is this possible?
Terrestrial Insects
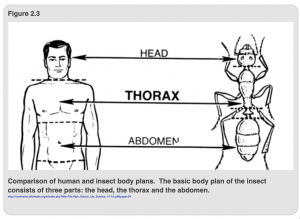
Unlike humans, where air enters only through our nose and mouth and is taken into our lungs, terrestrial insects have holes, called spiracles, in their thoracic and abdominal segments (Figure 2.3). Air enters the insect through valves in the spiracles which relax to open the spiracle and contract to close it. Figure 2.4a shows the path of air once it is inside the insect. Briefly, air moves through a series of tube-like structures called tracheae (trachea, singular) that are subdivided into progressively smaller tubes (Figure 2.4b), much like the way that arteries divide into veins in humans. Most tracheae are lined with a special type of cuticle, or exoskeleton, called taenidia, which fortifies the structure and prevents the trachea from being compressed, much like the thin spiral wires that wrap around the hoses in vacuum cleaners. Other trachea lack taenida and are enlarged into air sacs; these are common in flying insects and likely serve as pumps, forcing air back into other trachea and out of the spiracles.


Eventually, the tracheae terminate in a specialized cell, called a tracheole, that shares some similarities with our own capillaries and alveoli. Like capillaries in humans, tracheoles are in direct contact with the insect tissue that needs to be oxygenated. As in the alveoli in our lungs, the concentration of oxygen in the tracheoles is relatively high compared to the surrounding tissue, which means that oxygen diffuses down its concentration gradient out of the tracheoles and into the insect cells. Similarly, the concentration of carbon dioxide in the tissues is high because the cells comprising the tissue are actively producing it, and this causes the carbon dioxide to diffuse out of the insect cells and tissues and into the tracheoles. From there, the CO2 moves in to the trachea and through the spiracles to the outside world (Figure2.4a).
In small insects, simple diffusion is generally sufficient to oxygenate every cell in the insect body. However, this is one of the factors that limits the size of insects. There is a limit, after all, to how much biomass can be efficiently and reliably oxygenated using diffusion. This is not to say that large insects don’t have a few tricks. Indeed, large insects can augment the activity of diffusion by actively pumping air into their bodies. This is done by strategically opening some spiracles while closing others. At the same time, the insect uses abdominal muscles to alternately expand and contract the abdomen. The combination creates a sort of wave of air that flushes oxygen through the entire tracheal system. Still, unlike vertebrates, insects do not have any means to concentrate oxygen within their bodies, and the larger the insect, the greater the distance oxygen must diffuse before reaching an individual cell. These two factors (i.e. the inability to concentrate oxygen and the reliance on simple diffusion to distribute oxygen to each cell) ultimately limit the size of creatures who must conduct respiratory activities without the assistance of a circulatory system.
Aquatic Insects
Respiration in humans is basically a single, predictable process. Insects, however, have reconsidered the process of respiration many times, and each time those inventive insects have come up with new ways to serve the respiratory function and, as a consequence, to invade new and diverse environments. We’ve been talking about insect respiration as though it takes place exclusively in the terrestrial environment. And for many insects, respiration is indeed restricted to the terrestrial sphere. But, you’ve probably noticed that there are insects living in the water and they have respiratory issues that are very different from those that live exclusively on land. Even more interesting, there are insects that live in the aquatic world for part of their life cycle and the terrestrial world for the rest. How do they manage this?
One adaptation, of which insects have made liberal use, is gills for breathing in water. Immature damselflies (also known as naiads), have gills located at the terminal end of their abdomen (Figure 2.5). The predatory naiads live in water as immatures and breathe using gills that are an outgrowth of the tracheal system. The gills are moved in a fanning motion to maximize contact with oxygen in the water; this movement may also help the naiads to swim. In addition, the gills are covered by a thin layer of cuticle that is permeable to both oxygen and carbon dioxide. Gas exchange, thus, proceeds by diffusion from water into and out of the gills.

Another common fresh water aquatic species, the caddisfly, also has larval forms that are aquatic. Unlike in damselflies, the caddisfly gills can be located on the thorax or the abdomen. The coolest variant on the gill theme, however, has to be the dragonfly, which bears its gills internally—in its rectum. The immature dragonfly uses muscular contractions of its abdomen to force water into and out of its anus. In addition to serving the function of respiration in a rather creative way, the forceful expulsion of water from the dragonfly anus also gives it a way to move forward rapidly, which a human might call jet propulsion.
For the creative and resourceful insect, gills are not the only way to breathe in water. Far from it. Some insects, such as larval mosquitoes, have opted for the breathing tube mechanism which operates pretty much like a snorkel (Figure 2.6). The breathing tube, or siphon, is usually located at the end of the abdomen and pushes through the surface film of a body of water, often assisted by hairs that are coated with chemicals that help break the surface film. The hairs also help keep water out of the tube. The tube is, thus, in direct contact with air in the terrestrial world. Air is drawn into the breathing tube where it enters the tracheal system and circulates by diffusion within the body of the insect.


A very simple way to “go aquatic” that doesn’t involve the inconvenience of creating new structures is to simply trap a bubble of air and use it as a source of oxygen under the water (Figure 2.7). Insects that use bubble respiration may trap the bubble under their wing or they may have adaptations to protect the bubble, such as hydrophobic (i.e. water fearing/repelling) hairs. Either way, if the bubble is situated over a spiracle, the insect now has a way to breathe under water, at least for a limited period of time. Additionally, the chemistry of the components of air comes into play in interesting ways to extend the life of the bubble. How does this work?
The composition of air is roughly 21% oxygen, 79% nitrogen, and 0.03% carbon dioxide. However, the concentration of nitrogen in water is much lower. Obviously, the aquatic insect is interested in extracting the oxygen from its air bubble. But the presence of the other gases, particularly nitrogen, can assist by prolonging the lifespan of the bubble. In fact, the presence of nitrogen helps the bubble to act as a physical gill that can scavenge oxygen from the surrounding water, thereby allowing the “compressible gill” to deliver up to eight times more oxygen than what was present in the initial bubble. One reason for this is simple diffusion. As the insect withdraws oxygen from the bubble, the concentration of oxygen in the bubble decreases. Diffusion of oxygen into the bubble from the water will occur as the insect withdraws oxygen from the bubble.
The second important thing to note about the gaseous composition of the bubble relates to the relative solubility of each gas in water: nitrogen is much less soluble in water than is oxygen, while carbon dioxide is highly soluble in water. Thus, expelled carbon dioxide quickly moves out of the bubble, while oxygen moves in at a moderate pace, and nitrogen slowly diffuses out. The presence of nitrogen, which is much more concentrated in the bubble initially, effectively makes the bubble last longer than it otherwise would, allowing the insect to stay under water for a longer period of time. Ultimately, though, the size of the bubble decreases and the level of oxygen in the bubble falls to levels that are not sufficient to oxygenate the insect, and it will be forced to return to the surface to obtain another bubble.

Finally, another creative insect-based solution to aquatic life is to make a plastron. A plastron is a structure which, like the air bubble, acts as a physical gill. It is formed by hairs (Figure 2.8) or bumps on the surface of the insect cuticle that trap a thin layer of air, which the insect can use to fulfill its oxygen needs under water. Unlike the bubble/compressible gill, however, the size of the air pocket doesn’t change and therefore the insect does not need to return to the water surface to recharge the plastron. The interplay of the two major gases, oxygen and nitrogen are again important. As with the bubble, when oxygen is withdrawn by the respiratory activities of the insect, the lost oxygen can be replaced by removing oxygen from the water. And as was the case for the bubble, nitrogen will diffuse out of the plastron but at a slower rate than oxygen because of its lower solubility in water. This creates a partial pressure deficit in the plastron. Oxygen rushes in to correct the deficit created by the loss of nitrogen, thus giving the insect more oxygen to work with than afforded by the use of a bubble. Another way of looking at this is that the plastron replaces lost nitrogen with oxygen and this is a great benefit to the insect that needs oxygen under water to breathe. However, this mechanism of breathing isn’t compatible with high oxygen consumption and therefore the insects that have plastrons are much less active.
Insect Circulatory System
Perhaps you’ve noticed from the previous section that, unlike in humans, the circulatory system and the blood aren’t involved in gas exchange in insects. But, rest assured, insects do have a circulatory system. It is, however, quite a bit different from the human arrangement, both in terms of anatomy and function.

In insects, the heart or cardiac vessel is located in the abdomen. As can be seen in Figure 2.9, the heart may extend the length of the abdomen. Typically, an insect heart will beat somewhere between 30-100 times per minute. The main job of the heart is to send blood forward to the thorax through a tube called the aorta that extends from the heart to the head. By contracting, the heart that moves the blood forward. When blood reaches the end of the aorta in the thorax, the blood spills out and bathes the brain and muscles in the head. After providing nutrients and hormones to the head, the blood trickles back through the rest of the insect, eventually returning to the abdomen where it will re-enter the heart through special valves called ostia, whose function is to maintain blood flow in one direction.
Interestingly, the open circulatory of insects makes it difficult to get blood into certain places like antennae and legs. So, insects have fixed this problem by adding “pulsatile organs” that act like auxiliary mini-hearts to help circulate blood into those hard to reach areas. Sheer genius!
This very brief description of the insect circulatory system should have alerted you to several important differences in the circulatory systems of humans and insects. We’ve already discussed the fact that in humans there is an important collaboration between the respiratory and circulatory systems in oxygenating tissues. In insects, the circulatory system isn’t involved in gas exchange. You may have also noted that aside from the aorta, there are no blood vessels. Indeed, the insect circulatory system is considered to be an open circulatory system because the blood is not contained in vessels like it is in humans, but is in direct contact with internal organs in the body. Not surprisingly, this means that the function of the circulatory system is going to be somewhat different in insects and humans.
Some of the differences are manifest in the composition of the blood. Yes, insects do have blood, called hemolymph. However, unlike human blood, insect hemolymph lacks hemoglobin, the protein that binds oxygen and gives our blood its red color, and is not involved in delivering oxygen to individual cells. Thus insect hemolymph is generally not red, but is usually a yellowish or green color, which you may have seen after stepping on an insect. Interestingly, there are a small number of insect species such as midges (bloodworms that live in contaminated water) or horse bot flies (that spend most of their life cycles migrating through the tissues of the horse) that do have hemoglobin. These rare species of insects need the oxygen–binding capacity of hemoglobin to provide enough oxygen to live in hypoxic (hypo = low; oxic = oxygen) environments like polluted water and the insides of a horse. Because they have hemoglobin, their hemolymph is a reddish color.
Another difference in the composition of human and insect blood is that 90% of insect blood is liquid or plasma while only 10% consists of cells. In contrast, about 55% of human blood is plasma and 45% consists of cells. The plasma component of insect blood contains rather high levels of key nutrients (e.g. protein, sugars,and inorganic ions) in comparison to vertebrate blood. In addition, insect blood may contain particular sugars, hormones, and nutrients that have physiological and metabolic functions that are unique to insects. The sugar trehalose, for instance, is used by insects as an energy storage molecule but is not found in vertebrates. Instead, we use glucose as the major sugar in our blood. Similarly, insect blood may contain high levels of glycerol in the winter, which serves as a cryoprotectant or antifreeze to prevent the insect from freezing even when they are exposed to sub-zero temperatures.
Chapter Summary
We may have also left you with the impression that insect circulatory and respiratory systems are more “primitive” than those in humans. This, too, is a misconception. Multicellular life on earth actually began in the water. As noted in Chapter 1, the earliest arthropods were also aquatic, and insects evolved from such marine crustaceans. Early insects and other terrestrial arthropods were able to surmount the barriers to life on land, such as the development of respiratory systems that allowed them to extract oxygen from the air, and a cuticle that both served a protective function and prevented excessive water loss in a terrestrial world. More remarkable still, after becoming successful on land, several groups of insects then acquired the many adaptations we discussed in this chapter that allowed them to return to life in water, such as plastrons/compressible gills, as well as physical gills inside their rectums, on their abdomens and thoraxes. And as we mentioned in Chapter 1, insect wings likely evolved from thoracic gills, giving insects the opportunity to disperse even further and occupy even more unique habitats.
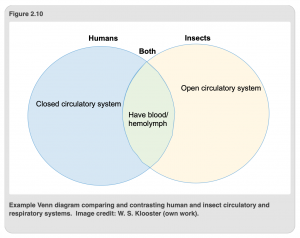
Entire volumes have been written both about the insect respiratory and circulatory systems. It is not our goal here to turn you into insect physiologists. However, this short introduction should be sufficient to acquaint you with the similarities and differences between the respiratory and circulatory systems in insects and humans. As a good review exercise/check for understanding, we encourage you to create a Venn diagram comparing and contrasting the circulatory and respiratory systems of humans and insects (see example: Figure 2.10). We will continue this approach in Chapter 3 with a discussion of the insect digestive and excretory systems.