Chapter 4 – Growth and Development

One of the regular features of animal life is that we grow and develop. No matter what form we are in when we start life, we will change over time, both structurally and biochemically. We nominally call this growth and development. Further, the sum of all the changes that take place over the course of animal life is referred to as the life cycle.
As you have all undergone growth and developmental processes yourselves, you are well-versed in the particulars of how your body has changed structurally over the course of your lifetime. But one issue that might require a little explication is what triggers the developmental changes that humans undergo in the course of “growing up.” From the health classes that you have likely taken in middle and high school, you probably also have a general understanding that hormones (chemicals that are produced by one organ, which then travel to another to have their effect) are the key factors that regulate our growth and development. Therefore, it is not surprising that the same is true of insects. However, the chemical structures and modes of action of insect hormones are, in many cases, quite different from those in humans.
In the case of both insects and humans, the master regulator of growth and development is the endocrine system. The endocrine system consists of a series of glands that secrete hormones that will circulate around the body to promote growth and development. This is usually mediated by having the hormone bind to receptors that are widely distributed around the body in order to elicit a specific effect. However, the role of the endocrine system in both insects and humans is as broad as it is important, and is not only limited to regulating growth and development. Indeed, another major function of the endocrine system is to maintain homeostasis (balance) within the body, as was discussed in Chapter 3. To this end, the endocrine system in humans and insects regulates appetite, metabolism, digestion, water balance, heart rate, and reproduction, among other things.
Rather than go into any lengthy detail of the myriad hormones or components of either the human or insect endocrine systems, in this chapter we focus specifically on a few of the key hormones that are involved in human and insect growth and development. As we have done previously, we will highlight both the similarities and differences between developmental processes and hormones in humans and insects. Moreover, we will illustrate the key experiments that were responsible for uncovering many of these discoveries in insects, and how we humans have capitalized on the differences in insect development to generate novel and effective pesticides. At the end of this chapter, you should therefore be able to compare and contrast the processes of growth and development in insects and humans.
Human Growth and Development
As a result of being well-endowed with hormones, humans progress through a very well-established series of changes as we grow from infant to adult. A reductionist view is that our bones, muscles, skin, internal organs, and veins must grow gradually. Generally, this growth continues until puberty and is mediated by many of the hormones and growth factors shown in Figure 4.1. The primary hormones that are responsible for human growth are growth hormone-releasing hormone (GHRH) and somatostatin. These stimulate and repress, respectively, the anterior pituitary gland from releasing the final growth hormone, human growth hormone (HGH). Both GHRH and somatostatin are produced by the hypothalamus (Figure 4.2).
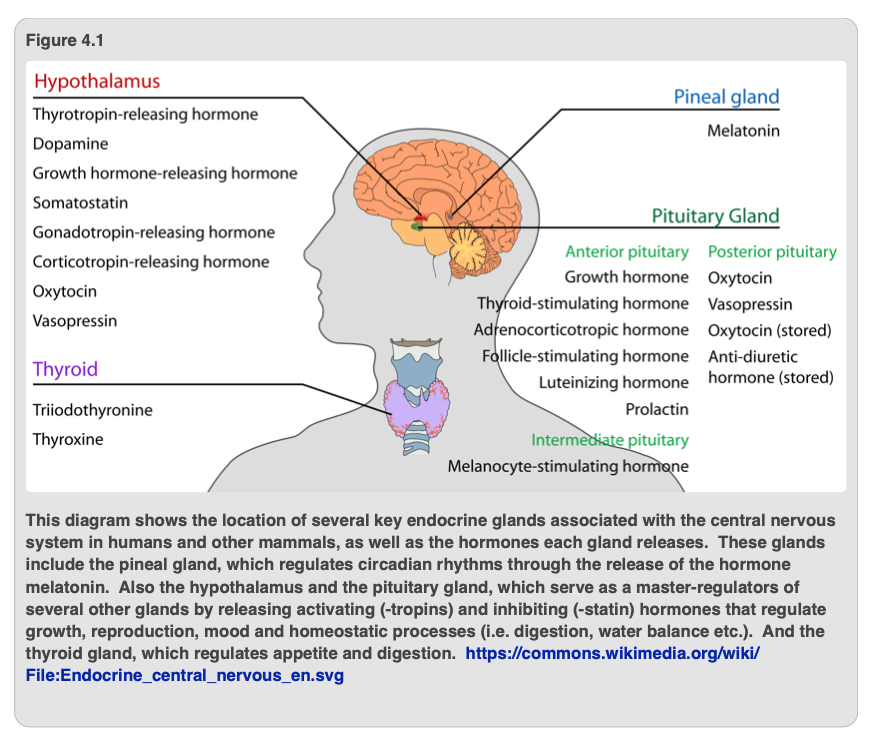

The proper regulation of all of our hormones is critical for normal growth and development. For example, Figure 4.2 illustrates what can happen if either too much HGH (gigantism) or too little HGH (dwarfism) is released from the pituitary gland. Even though our bodies stop growing in height (although not in girth) by the time we are adults, many of the hormones that induced growth in our earlier lives will continue to have activity in adulthood. The function, however, may change. For instance, HGH is still secreted in adults and has numerous additional roles. These include strengthening bones, increasing muscle mass and protein synthesis, maintaining proper levels of glucose in the blood, and stimulating the immune system.
Additional Hormones Responsible for Human Growth & Development
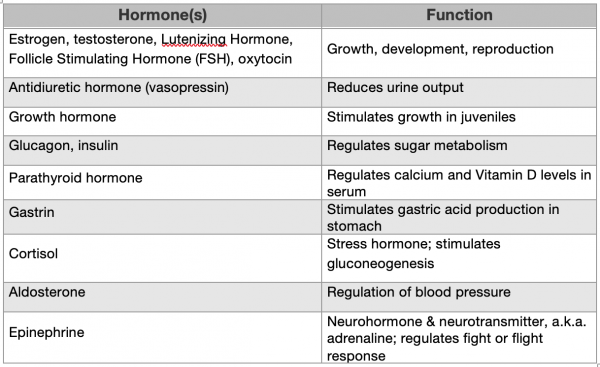
We have already discussed the three major hormones that are responsible for growth in humans (human growth-releasing hormone, somatostatin, and human growth hormone), and you are also likely familiar with several other hormones that we humans produce if only because you watch TV. You’ve most likely heard of estrogen and testosterone, which are two important hormones for the human reproductive cycle. However, there are several more hormones that regulate the growth, development, and reproductive processes in humans. In addition, others are involved in regulating homeostatic processes, such as water balance/urine output (vasopressin), digestion (gastrin), metabolism (parathyroid hormone, glucagon, and insulin), blood pressure (aldosterone), and mood (oxytocin, dopamine, and serotonin). Furthermore, as we saw with HGH, an individual hormone can have multiple effects. For example, epinephrine (a.k.a. adrenaline) increases heart rate and blood flow to muscles, elevates blood sugar levels, and causes your pupils to dilate, all of which are helpful if you need to fight or flee from a threat. See Table 4.1 for a summary of these human hormones and their function.
Table 4.1. Human hormones controlling growth, development, reproduction, & homeostasis (abbreviated list)
Insect Growth and Development
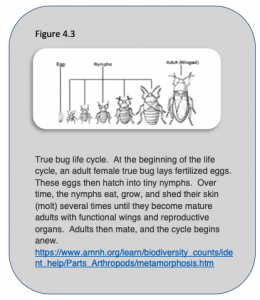
Given that insects and humans are both a type of animal, we would expect each of them to have mechanisms for growing and developing. In that respect, humans and animals are quite similar. However, this is one area in which the similarities between the two groups pale in comparison to the differences. The differences manifest immediately in that among insects, we find out that there is not one general life cycle, but two: complete and incomplete metamorphosis, which we will cover later in this chapter. But we also need to note that the changes that take place in the various parts of the life cycle are so extreme that we use a special term to describe the changes, i.e., metamorphosis, which is Greek for “change in form.” Incomplete metamorphosis (or hemimetabolous development), is the ancestral mode of development among insects and other arthropods, and is quite similar to the gradual development that characterizes most other animals. The life cycle begins with the egg (Figure 4.3) that hatches into an immature form of the insect, called a nymph, that looks like a miniature adult. There are several additional juvenile/nymphal stages that become successively larger as the insect continues to eat, grow, and undergo metamorphic events (molts) when its skin is shed. The stage of the insect between each successive molt is called the instar. As it molts, the nymph gradually develops its wings, such that the wings become larger and larger with each successive nymphal stage. However, the wings are not fully functional until the insect reaches adulthood. Furthermore, all immature forms lack functional gonads and cannot reproduce. Fortunately though, by the time an insect emerges as an adult, it can both fly and mate.
In contrast to the hemimetabolous life cycle, other insects undergo a radically different life cycle known as complete metamorphosis, also called holometabolous development (Figure 4.4). These insects, too, begin life as eggs. However, the eggs hatch into a larva (plural = larvae) that looks somewhat like a worm. Like the nymphs of hemimetabolous insects, the larvae eat, grow, and molt several times, such that each new larval stage is larger than the previous form. However, unlike hemimetabolous insects, the larval forms never resemble the adults, and lack compound eyes and wing buds. In a typical holometabolous insect, there are typically 3-5 larval instars. Eventually, the larva will molt into a pupa that looks quite different than either its earlier larval stage or its final adult stage. The pupa will undergo a final molt to become an adult insect that has both fully functional wings and gonads.
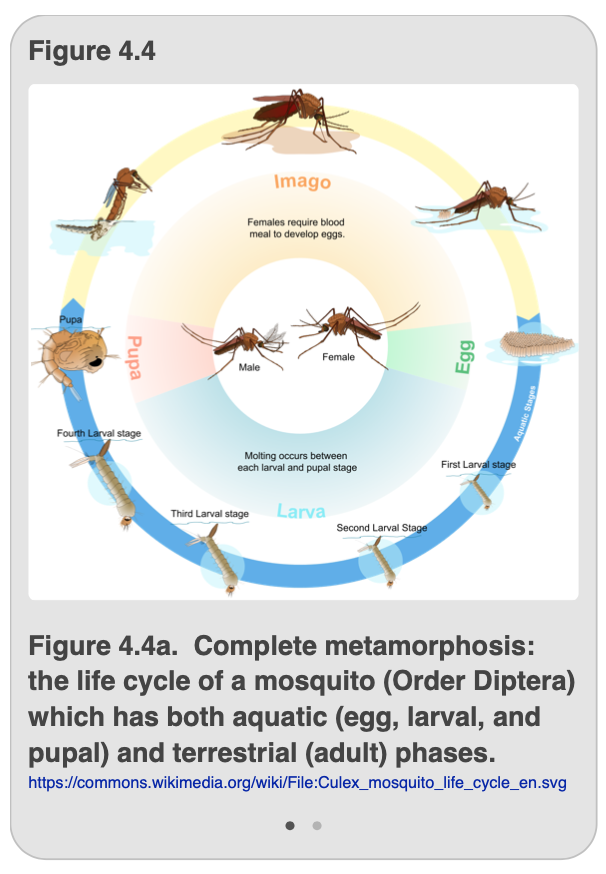

As you can see from Figure 4.4a and 4.4b, the different life stages can vary considerably in form despite the fact that both depict insects that undergo complete metamorphosis. Indeed, because the larvae of different holometabolous insects vary considerably, they are generally given unique names. For example, the larvae of butterflies and moths are called “caterpillars,” whereas those of beetles are often called “grubs,” and fly larvae are referred to (lovingly) as “maggots.” For both the mosquito and butterfly shown in Figure 4.4, the eggs hatch to form a series of larval instars followed by a very different looking pupa that undergoes a final molt to produce the adult. In the case of mosquitoes, we have the added complication that the egg, larval, and pupal forms are aquatic while the adult is terrestrial; in contrast, all life stages of the butterfly are terrestrial. Yet you can clearly see that for both mosquitoes and butterflies, the requirements of the holometabolous lifecycle are met: the immature forms look very different from the adults and lack the functioning wings, compound eyes, and external genitalia found in the adult forms. Also, a pupal form is present preceding the adult form. Table 4.2 summarizes the characteristics of incomplete and complete metamorphosis in insects.
Table 4.2. Characteristics of hemimetabolous and holometabolous metamorphosis
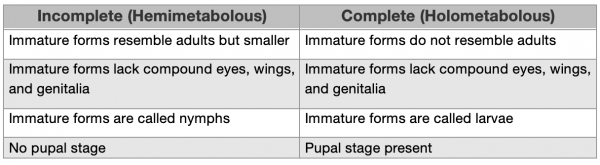
The Molting Process in Insects
At this point, you may be asking yourself: what exactly is this thing called molting? As we saw in the previous section, whether an insect undergoes hemimetabolous or holometabolous development, each insect molts several times throughout its life. Therefore you already know that molting is a necessary part of the insect life cycle and you probably have a sense that it might be important to metamorphosis. And indeed it is. But the first question you might want to have answered is why insects molt in the first place. And the basic answer to that question is that insects don’t have an internal skeleton, like humans do to prevent their bodies from collapsing into a heap. Instead, insects wear their skeleton on the outside of their bodies. It is called an exoskeleton for that reason. And if you’ve stepped on an insect and heard it go crunch, it does that because you have crushed its rigid exoskeleton. Figure 4.5 shows all of the many layers of the insect exoskeleton. The level of detail might seem overwhelming, but the key points here are that the three major layers of the insect exoskeleton (the epicuticle, exocuticle, and endocuticle), are not living tissues. Instead, the exoskeleton is more similar to our hair and fingernails in that it is secreted by living epidermal cells, and is coated with waxes and other secretions that are produced in glandular cells and moved through pores or channels.
The thickness and toughness of the exoskeleton varies among different insect species; for example, an adult rhinoceros beetle is much harder than an adult flesh fly. Exoskeletons also vary throughout an insect’s development (beetle grubs are less crunchy than adult beetles), and even among different parts of the insect (the heads of caterpillars are much harder than their thoraxes and abdomens). However, in all cases, the exoskeleton offers the insect protection from the environment, provides a structure to which they can attach their muscles, and can be coated in waxes that allow insects to conserve water.
In spite of the manifold benefits the insect exoskeleton offers, it imposes a very important constraint: once the exoskeleton is formed, it cannot expand. Therefore, in order to proceed through its life cycle and grow, an insect must escape from its old, small exoskeleton and form new, larger one. This is the rationale behind the whole metamorphic scheme.
This begs the question: what exactly does the molt involve? From our descriptions so far, you might correctly guess that the first thing that has to happen is the separation from the old exoskeleton. In a process called apolysis, the endocuticle (the innermost layer of the exoskeleton) separates from the underlying epidermal cells. A whole bunch of things will take place after the cuticle has been detached but before it is formally shed. It is kind of like putting on a new set of clothes while you’re still wearing the old set.
The next step is that those same epidermal cells will secrete molting fluid that will eventually digest the old endocuticle. However, it is initially released in an inactive form. The third step is to generate a new cuticulin layer. The cuticulin layer is composed of lipoproteins (part lipid/fat and part protein) that protects the insect from the digestive activity of the molting fluid. Functionally, the cuticulin layer will become part of the new epicuticle—the thinnest and outermost layer of the exoskeleton. Now things really heat up. The molting fluid that was initially inactive is activated. It has powerful enzymes that literally digest the old endocuticle, which is the innermost layer of the exoskeleton. As the old endocuticle is broken down, the products of that digestion pass through the cuticulin layer where the waiting epidermal cells will recycle them.
Insects are incredibly efficient at conserving their resources, and as much as 90% of the old endocuticle will be recycled through this process. This is why the cuticulin layer is so important. The insect needs to not only protect the epidermal cells from the powerful enzymes but also must differentiate between the old cuticle it wants to replace and the new cuticle it wants to keep.
After the old endocuticle has been digested, the epidermal cells secrete a new undifferentiated procuticle (future exocuticle and endocuticle). Since insect muscles are anchored in the insect body by attaching the muscles to the exoskeleton, the muscles have to be detached from the old cuticle that is about to be shed and reattached to the new exoskeleton. After the maximum amount of old endocuticle has been recycled, new procuticle has been synthesized, and the muscles are appropriately attached to the new exoskeleton, then the molting insect takes in a lot of air causing the old exoskeleton to split along a line of weakness, called the ecdysial sutures. Emergence from the old cuticle, a process officially called ecdysis (Greek for “get out!”), has now occurred.
Keep in mind, there may be major structural changes going on in the insect while it molts and moves from one life stage to the next. Recall, for instance, that the physical form of a larva is much different than a pupa, which is much different from the adult form. In the next section we will discuss the specific hormones that signal to the epidermal cells what kind of exoskeleton to produce (i.e. larval/nymphal, pupal, or adult). However, the complexity of this entire process is one of the things that makes metamorphosis so amazing (see Table 4.3).
Table 4.3. Steps involved in metamorphosis in insects

Despite having shed the old cuticle, the exoskeleton of the newly emerged insect is still in progress. Once liberated from the old cuticle, the insect continues to take in air so that it can fully expand into its new, larger exoskeleton (sort of like inflating a balloon). The new cuticle will be very soft and flexible, thereby allowing for its rapid expansion. The new cuticle will also initially be light in color, generally white, even if the insect is ultimately darkly colored (Figure 4.6). The insect is extremely vulnerable during this stage because it is not protected by a hard shell, nor does it blend in as well with its surroundings.

However, insects do not stay in this vulnerable state for very long, and the new cuticle will proceed to undergo hardening and darkening processes within a couple of hours after the molt. Hardening strengthens the exocuticle by cross-linking chitin (the major component of the insect exoskeleton) with proteins by joining them to quinone molecules. Darkening results from the formation of pigments, which impart the normal color of the insect. The process of metamorphosis is now finished—until the insect undergoes its next molt. Follow the link (Video 4.1) to watch a excellent Youtube video produced by Dr. Larry Keeley at Texas A&M that beautifully illustrates and summarizes the molting process in insects.
Hormones Responsible for Insect Growth & Development
Similar to humans, hormones are responsible for growth and development in insects. However, in insects, the two main hormones that trigger developmental changes are called ecdysone, a.k.a., the molting hormone, and juvenile hormone (JH). The function of ecdysone and JH are similar to HGH and estrogen/testosterone in humans in that they trigger development and key elements of subsequent reproductive events. However, the function of the insect hormones can only be understood by considering the action of both ecdysone and JH together.
In Figure 4.7, you see the stages of the life cycle of a holometabolous insect detailed on the X-axis, while the titer (concentration) of ecdysone and JH are shown on the Y-axis. As previously mentioned, ecdysone and JH work in concert to determine the outcome of a molt. In other words, metamorphosis, is dependent on the interaction of the two hormones. Ecdysone is also known as “molting hormone,” which suggests that the presence of large amounts of ecdysone triggers a molt. Therefore, you will notice that a glut of ecdysone is released immediately prior to any molt. Indeed, this hormone triggers apolysis, which you will recall is the separation of the epidermal cells from the old exoskeleton to begin the molting process. Figure 4.7 is actually a simplified graph that shows the amount of ecdysone as the same at each molt, but in reality the amount of ecdysone will vary from one molt to the next. However, there is a reliable spike of ecdysone released slightly before each molt, which signals to the insect to undertake all of the activities necessary to accomplish the molt.
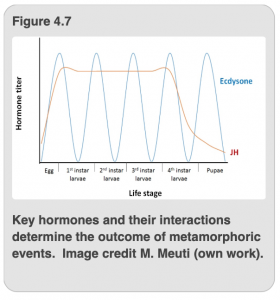
As we discussed in the previous section, the epidermal cells require a signal to separate from the old exoskeleton, as well as to determine what kind of new cuticle they should produce (i.e. either nymphal/larval, pupal or adult structures). The outcome of each molt is determined not only by ecdysone, but also the level of JH at the time of each molt. Remember, JH stands for juvenile hormone, a descriptor that indicates its function. If JH is present in high amounts when the signal to molt is queued by an upsurge of ecdysone, the juvenilizing effect of JH will result in another juvenile form (i.e. a nymph for hemimetabolous insects or a larva in holometabolous insects) as the outcome of that molt. During the last larval stage, the level of JH drops precipitously (Figure 4.7). Thus, when there is a surge of ecdysone but little-to-no JH is present, the result of that molt is not another nymphal stage, but the adult stage for hemimetabolous insects. Similarly, in holometabolous insects, when ecdysone peaks in the absence of JH, the insect molts into the pupal stage. The level of JH does increase during the pupal stage, where it has different functions, but then it drops again prior to the molt to adult form.
You might be wondering how anyone ever figured out the roles of ecdysone and JH in insect development (or if you weren’t, hopefully you are now!). The role of each of these hormones was uncovered by a very famous British entomologist named Sir Vincent Wigglesworth, using some very interesting and unconventional experiments. He worked with the South American kissing bug, Rhodnius prolixus, which is a hemimetabolous insect (Figure 4.8a, Figure 4.8b and Figure 4.8c). This species feeds exclusively on human blood, and requires a blood meal in order to molt from one phase to the next. Doing something that only entomologists can do (as opposed to people who work with other types of animals), Wigglesworth decapitated (removed the head) of a 5th instar nymph after it had taken a blood meal and was ready to molt into an adult, and attached it to a decapitated 1st instar nymph that had not blood fed and was not ready to molt. That’s right folks, he made Franken-bugs! Technically this is called parabiosis (the anatomical joining of two animals together), and it allowed the insects to transfer hormones and other factors in their hemolymph/blood to one another. Not only did both the 1st instar and 5th instar nymphs survive this process, but both insects successfully molted into adults; although the first instar nymph became a tiny, precocious, and (unfortunately) headless adult.


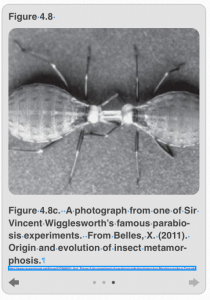
Why did this happen? Well, Wigglesworth correctly surmised that the high level of a molting hormone (which we now know is ecdysone) was produced by the 5th-instar nymph and transferred to the first instar nymph, causing both to molt. But why did they molt into adults? To determine this, Wigglesworth later transplanted different insect endocrine glands from 4th instar nymphs into blood-fed, 5th instar nymphs. He observed that when one particular gland, the corporus allatum, was transplanted into 5th instar nymphs, these insect underwent a supernumerary molt, becoming an extra-large, 6th instar nymph—something which never happens in nature. Therefore, he concluded that the corpora allata produced a “juvenilizing hormone.” Wigglesworth, yet again, correctly concluded that whereas the 1st instar nymph in his original parabiosis experiments were producing JH in addition to ecdysone, the level of JH became too dilute to exert its juvenilizing effect when it was spread between the large body of the 5th instar nymph and its own. Hence this is why both individuals were able to molt into adults.
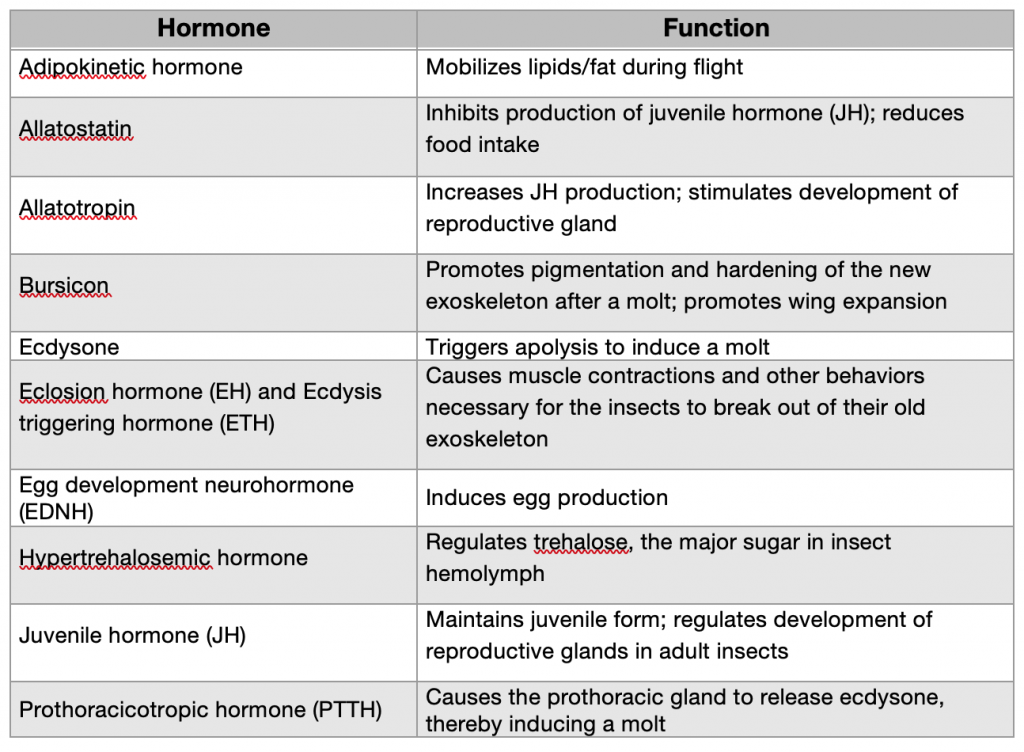
Of course, JH and ecdysone are not the only hormones found in insects, just as HGH and somatostatin are not the only hormones found in humans. Table 4.4 provides a partial list of insect hormones. You probably won’t recognize many of these since they control aspects of physiology that are unique to arthropods— such as promoting tanning of the newly formed exoskeleton (bursicon) or mobilizing lipids for flight (adipokientic horomone). However, be assured that, like your typical human teenager, insects are swimming in hormones.
Table 4.4. Insect hormones and their functions (abbreviated list)
Significance of Insect Growth and Development
Hopefully you are now somewhat amazed by both the complexity and sophistication of the insect endocrine system and its tight regulation of insect growth and development. Yet you may be wondering why any person would ever care about how or why insects are able to regulate their growth. As it turns out, humans have discovered ways to manipulate the differences between our own development and those of insects for the purposes of pest control. For example, the chemical methoprene is very similar in structure to JH, and when it is applied to insects it causes them (much like in Wigglesworth’s experiments) to undergo an extra nymphal or larval stage. This often has deleterious effects on the adult insects, and hence methoprene is an effective pesticide. Furthermore, methoprene is applied at low doses to late instar silkworm caterpillars. Can you imagine why? Well, since methoprene mimics the effects of JH and causes the caterpillars to undergo an additional larval molt, this leads to extra-large caterpillars and therefore to extra-large pupae and silk cocoons, and hence increased silk production.
Also of interest, there are times when insects must slow down or arrest their development, such as during extended periods of time when food might be limited or temperatures might be too low for the insects to move around. Indeed, most temperate insects enter such arrested periods of development, called diapause during winter, making it similar to mammalian hibernation. Other insects that live in warmer, tropical climates may similarly aestivate, which is to say they enter a “summer diapause” in order to survive in periods when water or food might be limited. These periods of arrested development are controlled by the very same hormones that we have reviewed here. Several researchers (including our very own Dr. Meuti!) are working to determine how we can manipulate insect growth and development to prevent insects from appropriately timing their entry into and exit out of diapause, and hence causing them to commit ecological suicide. In case you are interested, here is a link to a video of Dr. Meuti discussing her research on this topic.
Chapter Summary
Just like humans, insects grow and develop over time, yet the insect life cycle is quite different from that in humans. Insect lives are punctuated by molts, when the insects must shed their old, small exoskeletons and create new, larger ones. The molting process is quite complex, but the type and timing of molt is regulated by a few critical hormones. Just as we saw that elevated levels of HGH cause gigantism whereas low levels lead to dwarfism in humans, manipulating the levels of JH in insects can have severe consequences for insect growth and development. As you continue to reflect on the similarities and differences between the endocrine systems as well as growth and development in insects and humans, we hope that you gain a fuller appreciation of the miracle of life and all of the various changes that both you and the fly on your wall underwent to successfully reach adulthood.