Chapter 12: Male Reproductive System
The testes
The testes have two functions. The first is to produce spermatozoa and the second is to function as an endocrine organ. A portion of mesonephric cells develop into cords of tissue called seminiferous cords which incorporate the migrating primitive germ cells into their structure. The seminiferous cords, also known as medullary cords, develop eventually into Sertoli cells, while mesodermal cells in between seminiferous cords develop into Leydig cells (interstitial cells). Sertoli cells secrete factors that regulated spermatogenesis and spermiogenesis, the function of Leydig cells. They also secrete tubular fluid and produce inhibin that regulates hormone production. Leydig cells are responsible for testosterone production.
Spematogenesis
Germ cells residing in the developing testes are referred to as spermatogonia. Successive rounds of mitotic division increase the number of cells that can end as the terminally differentiated spermatogonia, known as spermatozoa. Spermatogenesis is quiescent until puberty and the process takes place within the seminiferous tubules.
The spermatogenic process can be divided into the mitotic, meiotic and metamorphic periods. Spermatogenesis takes places at different stages throughout the length of the seminiferous tubules.
The mitotic phase of spermatogenesis results in increased numbers of spermatogonia, allowing for an enlarged replicative pool before the first meiotic division. Two types of spermatogonia are present. Type A spermatogonia have a large round nucleus with condensed chromatin, peripheral nucleoli and a nuclear vacuole occasionally. These spermatogonia maintain the replicative pool of germ cells. Type B spermatogonia have dispersed chromatin, central nucleoli and no nuclear vacuole. These spermatogonia eventually produce spermatozoa.
Formation of spermatocytes represents the meiotic stage of spermatogenesis. Primary spermatocytes are derived from the spermatogonium and are still diploid cells. Primary spermatocytes undergo the first meiotic division to become haploid cells called secondary spermatocytes. These soon enter the second meiotic division to become spermatids.
Spermiogenesis
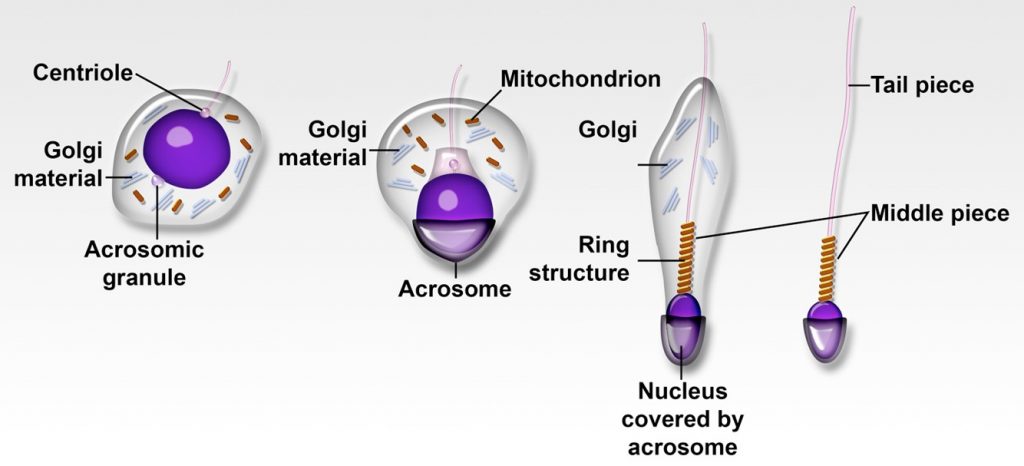
The conversion of spermatids to spermatozoa represents the metamorphic stage known as spermiogenesis. During this process, granules in the Golgi vesicles coalesce to form a large vesicle (acrosomal vesicle) containing a structure called the acrosome. The acrosomal vesicle expands to cover half of the nucleus. The acrosome and the acrosomal vesicle form the (acrosomal) head cap. Simultaneously, centrioles, mitochondria and the Golgi remnants migrate to the cytoplasm opposite to the acrosomal cap. One centriole elongates to become the developing flagella. The centrioles also interface with the nucleus to form the neck. During the entire process, the spermatid elongates and it nucleus condenses.
The mature spermatozoon is divided into 5 sections
- Head
- Neck – where the centriole migrated and connects the head to the tail
- Midpiece
- Principal piece
- Tail or end piece
The stage of spermatozoon development is variable between regions of seminiferous tubules and progression is cyclical in nature. This variability is known as the spermatogenic wave.
The activity in the seminiferous tubules is primarily regulated by the products of the hypothalamus and the anterior pituitary. Luteinizing hormone (LH) and Follicle stimulating hormone (FSH) are secreted by the anterior pituitary. FSH initiates spermatogenesis, while LH stimulates testosterone secretion that is required for spermatozoon maturation.
Testicular structure
A capsule composed of dense collagenous material that encloses the testicle is called the tunica albuginea. Most species contain a vascular layer within the tunica albuginea. Horses often contain smooth muscle fibers within the tunica albuginea. The double layer of simple squamous epithelium and mesentery directly covering the testes and apposed to the tunica albuginea is the tunica vaginalis.
The parenchyma of the testicle is divided into lobules by loose connective tissue bands (septuli testes). These lobules are composed of tubules lined by stratified epithelium composed of maturing germ cells (spermatogonia, spermatocytes, spermatids and spermatozoa) and Sertoli cells. These tubules are supported by a basement membrane which contains fibroblasts and myofibroblasts. The Sertoli cells are triangular or oval in shape with a prominent nucleolus and fine chromatin. These tubules (seminiferous tubules) constitute the exocrine portion of the testes. The maturing germ cells are stratified so that the spermatogonia constitute the basilar compartments closest to the basement membrane. The spermatocytes, spermatids and spermatozoa constitute the next layer or the apical compartment with the spermatocytes adjacent to the basilar compartment and the spermatozoa closest to the lumen.
The endocrine portion of the testes is comprised by the Leydig or interstitial cells which are present between seminiferous tubules primarily located in the extra-tubular connective tissue. These cells are responsible for testosterone secretion.
As spermatozoa mature in the seminiferous tubules, they are guided out of the testes by the rete testis (randomly arranged) or tubuli recti (straight tubules) which are connected to the seminiferous tubules. These tubules eventually lead to the extragonadal tubular structures.
