Chapter 5: Bone
Bone growth
Bone can grow by two mechanisms: membranous ossification and endochondral ossification. Membranous ossification occurs primarily in flat bones (ie. bones of the skull, mandible), while endochondral ossification is seen in long bones (ie. appendicular skeleton).
Membranous ossification
Membranous ossification begins as condensation of primitive mesenchyme, and cells differentiate into an osteoblastic phenotype. These cells form bone directly that is laid in apposition, on top of the bone that has just formed. Importantly, there is no cartilage intermediate. Membranous ossification is also the way in which bones (including long bones) grow in width. It therefore contributes a significant portion to the diaphyseal cortex of long bones.
Endochondral ossification
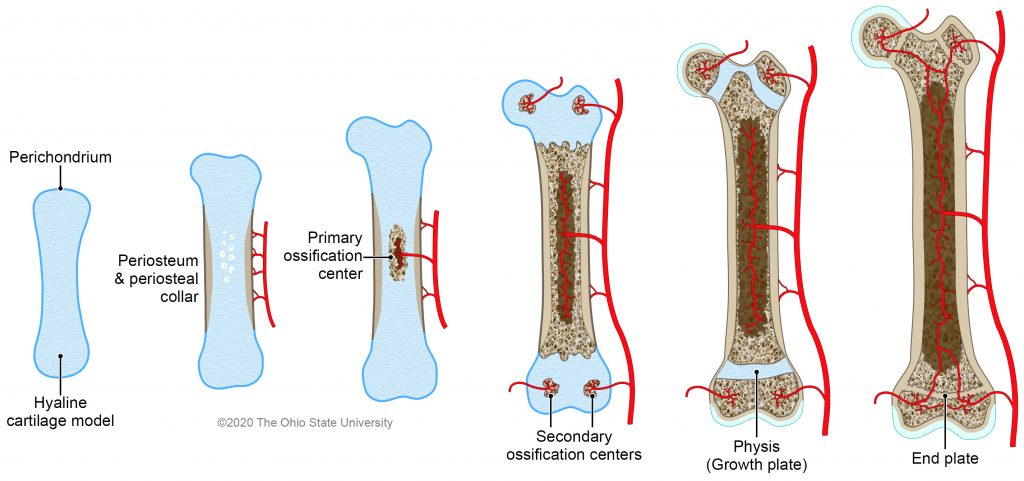
Endochondral ossification significantly differs from membranous ossification. A cartilage model that becomes almost entirely replaced by bone precedes the formation of the actual bone. Increases in bone length are primarily the result of endochondral ossification, as the cartilage model can grow interstitially (from within the matrix). In the developing fetus, primitive mesenchyme condenses and these cells differentiate into a chondroblastic phenotype. The chondroblastic population proliferates to form a cartilage model that approximately mimics the final bone contour. Because this model is initially composed entirely of cartilage (considerably less rigid than bone), the developing bone can grow interstitially. Initially, the perichondrium that surrounds the cartilage model becomes invaded by blood vessels, forming periosteal collar around the diaphyseal region. With this periosteum, there are osteogenic precursors that form an bony sleeve around the primary center of ossification. This primary center of ossification begins in the middle of a long bone as blood vessels invade into the cartilage model. With time, vessels then invade the periphery of the bone, establishing secondary centers of ossification. The primary and secondary centers of ossification progressively expand with endochondral ossification occurring at the edges of an ossification center. Once the ossification centers get to a certain point, there becomes a thin line of growing cartilage between the primary and secondary ossification center. This is known as the physis (‘growth plate’) and its primary function is to lengthen the bone. Endochondral ossification that occurs at the edge of a secondary ossification center, immediately beneath an articular surface, is called the articular-epiphyseal complex (AEC). Histologically the process of endochondral ossification is almost identical. Endochondral ossification in any given ossification center occurs at different rates at different loci. This allows the bone to form its shape and spatial alignment. Once endochondral ossification finishes at the AEC, the subchondral bone condenses to form an end plate.
The process of endochondral ossification can be seen in histologic sections of a growth plate (physis), where multiple distinct layers can be identified. These include the zone of resting chondrocytes, zone of proliferation, zone of hypertrophy, zone of mineralization, and zone of ossification. Chondrocytes within the zone of proliferation and hypertrophy are arranged into linear columns interspersed by chondroid matrix. For endochondral ossification to occur normally, three things must happen in a very orderly fashion. These are mineralization of the cartilage matrix surrounding the chondrocyte, death of the chondrocyte (mineralization stops transfer of nutrients causing the cell to die), and vascular ingrowth. Vessels grow between spicules of mineralized matrix. This process is particularly important as the new vessels bring osteogenic cells (osteoblasts) that can then deposit osteoid on top of the mineralized cartilage. The area containing mineralized cartilage spicules and early ossification is referred to as the primary spongiosa. In the secondary spongiosa, the newly formed bony trabeculae become modeled into mature trabeculae that reflect tension and compression on the bone.
